
One of the core principles of a Paleo diet is avoiding foods that contain compounds called toxic lectins. The term toxic lectin originates from the fact that lectins are a class of carbohydrate-binding proteins found in all foods (and indeed all forms of life, including humans!), but that only a subset of these are an issue for human health via food consumption (hence lectins versus toxic lectins). The lectins that are of concern have a few important properties: they are relatively stable through food preparation; they are difficult for humans to digest; they are known to interact strongly with the brush border of the intestine (which may impact cell viability and/or barrier function in addition to allowing protected transport of the toxic lectin into the body); and they are biologically active once they enter the body. There are two main classes of these toxic lectins: prolamins and agglutinins. Because gluten is a prolamin, their effects are typically more widely understood (and were discussed in detail in How Gluten (and other Prolamins) Damage the Gut). But, agglutinins are arguably more problematic for health, so let’s discuss them in detail.
Table of Contents[Hide][Show]
Note: this is a fairly technical article. I’ve tried to distill the most important information into “take-home messages” throughout. If you’re keen to get into the nitty gritty detail, I recommend reading the following which covers a lot of the background cell biology referred to in this post: How Gluten (and other Prolamins) Damage the Gut), Why Grains Are Bad, Part 1, Lectins and the Gut , What Is A Leaky Gut? (And How Can It Cause So Many Health Issues?), What Is the Gut Microbiome? And Why Should We Care About It?
What are Agglutinins?
Agglutinins are proteins characterized by their ability to induce the clumping (or agglutination) of red blood cells. Some agglutinins, such as ricin, which comes from castor bean casings, are so toxic that as little as one milligram, if inhaled or injected (presumably accidentally) intravenously or intramuscularly, is deadly. How much is one milligram of ricin? Roughly the size of a grain of sand. In fact, the Bulgarian dissident Georgi Markov was assassinated in London in 1978 with a pellet containing 0.2 milligrams of ricin that was shot or injected into his thigh, allegedly with the tip of an umbrella.
Agglutinins are part of the seed’s natural defense mechanisms, and while all the roles these lectins play in protecting the seed have yet to be discovered, what we do know is that they protect the seed from fungal infection and perhaps from insect predation as well. In fact, genetically modified grains typically contain higher levels of agglutinins to help protect industrial crops from pests. Wheat germ agglutinin (WGA) is so powerful at resisting insects that the gene for WGA has even been added to genetically modified corn.
Guide to Nutrivore e-book
Nutrivore is the simple yet revolutionary concept:
Choose foods to meet the body’s nutritional needs!
I’m very excited about the Nutrivore information! Thank you so much for all your research and hard work you put into this kind of information for all of us to use!!
Debbie

As is the case with prolamins (again, see How Gluten (and other Prolamins) Damage the Gut), agglutinins are difficult to digest because we lack proteolytic enzymes (digestive enzymes that lyse, or break apart, proteins) capable of breaking it apart into individual amino acids. It is also very stable at high temperatures and low pH, so neither cooking nor our stomach acid are much help in breaking it down, either.
Take-home message: agglutinins are proteins founds in grains, pseudograins, legumes, and vegetables of the nightshade family that can be toxic in high quantities. In low quantities, they are inflammatory and cause a leaky gut.
Wheat Germ Agglutinin
Wheat germ agglutinin (WGA) is the best-studied agglutinin, and probably also the most detrimental (although soybean agglutinin is a close second; we’ll get to that in a minute). WGA is not directly toxic to gut enterocytes (see What Is A Leaky Gut? (And How Can It Cause So Many Health Issues?) to review how these cells form the gut barrier) in concentrations equivalent to those normally found in food, but because our bodies can’t digest it, WGA travels through the small intestine largely intact. WGA is very good at interacting with the brush border of the intestine, and in doing so, increases intestinal permeability. (Hello, leaky gut.) It is also a well-known stimulator of the immune system. In fact, its activities in the gut qualify it as a biologically active protein, which is not normal (or generally healthful!) for proteins in our food.
How does WGA interact with the gut barrier?
The outer membrane of every cell is composed of a double layer of fat molecules (called the lipid bylayer) with various proteins (receptors, for one) that perform a variety of functions embedded within the bylayer. Some of the fats and proteins within the membrane have sugar molecules (which are important for a variety of normal membrane functions) attached to them on the outside-of-the-cell side of the membrane. These are called membrane carbohydrates, and they actually constitute 2–10% of a cell’s membrane. These membrane carbohydrates are one of eight different sugars: glucose, galactose, mannose, fucose, xylose, N-acetylgalactosamine, N-acetylglucosamine, and N-acetylneuraminic acid.

Nutrivore Weekly Serving Matrix
An easy-to-use and flexible weekly checklist
to help you maximize nutrient-density.
The Weekly Serving Matrix is very helpful! I’ve been eating along these lines but this really helps me know where to focus vs. which foods serve a more secondary role. It’s super helpful and has taken a lot of worry out of my meal planning. Thanks!
Jan

WGA binds specifically to two sugars (N-acetylglucosamine and N-acetylneuraminic acid) that happen to be membrane carbohydrates in all animal cells and form key parts of the glycocalyx of gastrointestinal cells (see What Is A Leaky Gut? (And How Can It Cause So Many Health Issues?)). WGA binds at a very high rate to human intestinal cells via these membrane carbohydrates, and WGA is rapidly internalized into the enterocytes, probably via endocytosis (see How Gluten (and other Prolamins) Damage the Gut).
One of the membrane glycoproteins that WGA binds to (by binding to the carbohydrate component of the glycoprotein) is the epidermal growth factor (EGF) receptor. The EGF receptor is well-known to promote what is called receptor-mediated endocytosis and may be the reason WGA is so readily internalized by gut enterocytes. Even more important, though, the EGF receptor regulates the paracellular permeability of epithelial cells—this means that when the EGF receptor is activated (for example, by binding with WGA), signals are sent throughout the cell that result in the opening of the tight junctions between enterocytes (analogous to How Gluten (and other Prolamins) Damage the Gut). It is thought that the EGF receptor plays an important physiological role in maintaining epithelial-tissue organization and permeability, thereby maintaining the integrity of the gut barrier.
Very low concentrations of WGA (but very possible to achieve from dietary sources) increase epithelial-cell permeability, meaning that WGA causes the tight junctions between epithelial cells to open up sufficiently for molecules to cross (at least scientists have measured this in isolated cells studied in the lab). WGA accumulates in lysosomes (whether this damages the lysosomes, as in the case of gliadin, has not been studied; again, see How Gluten (and other Prolamins) Damage the Gut)), and a fraction of the WGA crosses the epithelial barrier intact (though we don’t know how yet). And while the amount of WGA that crosses the epithelial barrier is low, it’s certainly enough to have a huge impact on the immune system. In fact, even at these very low concentrations, WGA stimulates the secretion of proinflammatory cytokines (chemical messengers of inflammation).
The Nutrivore Collection
Save $10 on Guide to Nutrivore and the Nutrivore Weekly Serving Matrix!
I never realized how important nutrients are and how intricately the body works! I can’t thank you enough for sharing all your knowledge and insights.
Cheryl

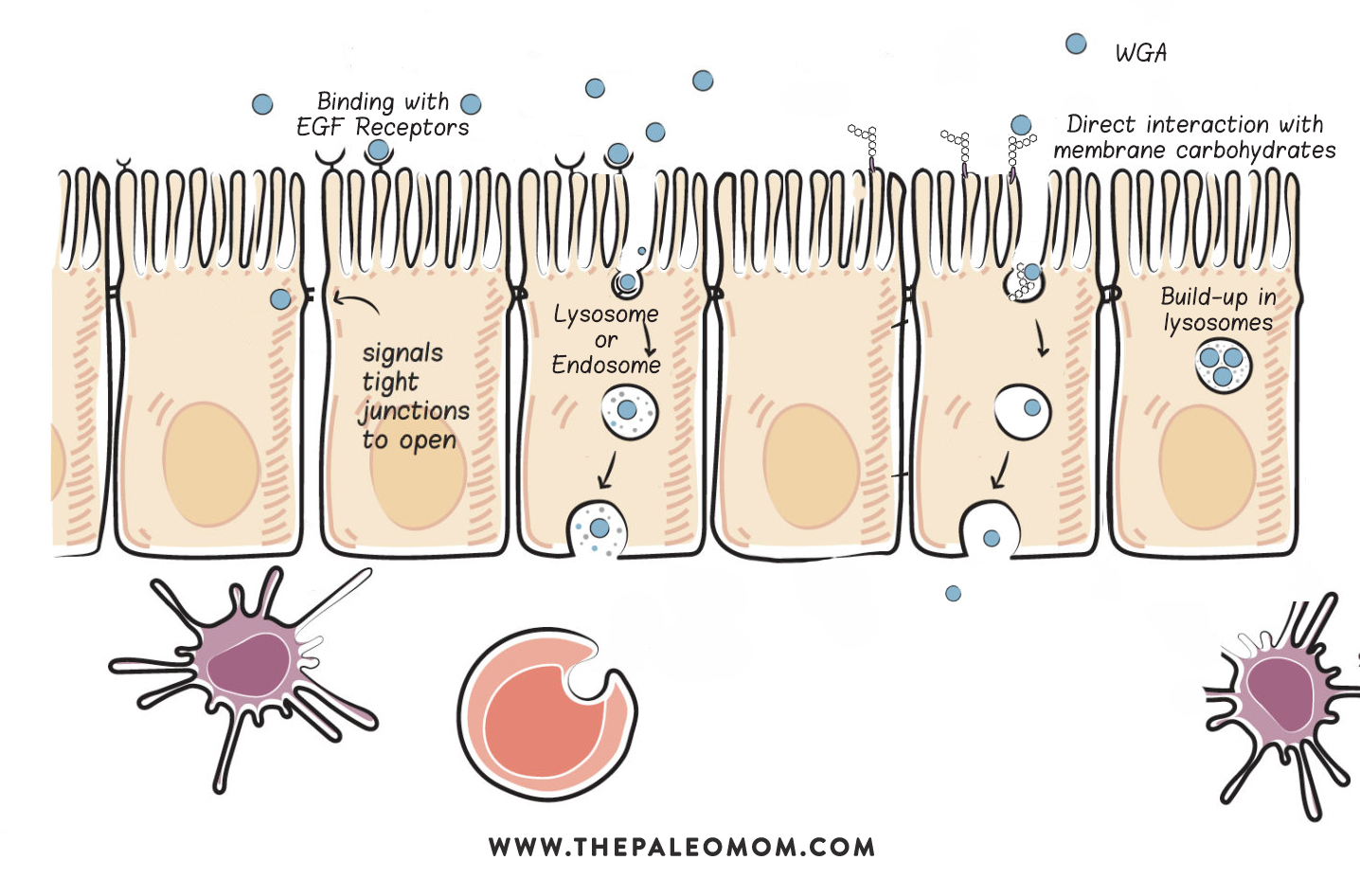
WGA is known to have a profound effect on both the innate and the adaptive immune systems. Some of the proinflammatory cytokines induced by WGA cause inflammation, including stimulating phagocytosis by neutrophils (“eating” of stuff by white blood cells) and stimulating the production of reactive oxygen species (oxidants). And once again, this generalized inflammation can cause damage to the innocent-bystander enterocytes. WGA also induces cytokine production known to increase proliferation (cell division) of all helper T cells. However, WGA can also bind to cytokine receptors on T cells and thereby inhibit proliferation (which may actually contribute to overstimulation of one subset of T cells, as is often seen in autoimmune and immune-related disorders; see The Autoimmune Protocol). WGA also stimulates the production of antibodies by B cells. And WGA binds to blood-derived immune cells, increasing activation of inflammatory cells and also increasing apoptosis of white blood cells.
WGA is so good at getting into gut enterocytes that it is being investigated as a carrier for oral drugs, the idea being that WGA could be bound to a drug molecule and help facilitate the drug’s absorption into the body. Fortunately, a flag has been raised about the possible toxic effects of WGA as a drug-delivery vehicle, with research suggesting that WGA may be the cause of various gastrointestinal disorders.
Take-home message: wheat germ agglutinin tricks the cells that line our digestive track into internalizing it and transporting it into the body. A side effect of this transport is increased intestinal permeability, aka leaky gut.
What About other Agglutinins?
 So what about other agglutinins? While most have not been as extensively studied as WGA, agglutinins from other plants have been shown to have similar effects.
So what about other agglutinins? While most have not been as extensively studied as WGA, agglutinins from other plants have been shown to have similar effects.
Kidney beans contain a high level of a very toxic and immunogenic agglutinin called phytohaemagglutinin (also sometimes called kidney bean lectin). Phytohaemagglutinin is also found to a lesser extent in cannellini beans, common beans, and broad beans, such as fava beans. As few as five raw kidney beans can cause extreme gastrointestinal distress, with symptoms like those of food poisoning (not that anyone would want to eat raw kidney beans). While soaking and then cooking beans greatly reduces the activity of phytohaemagglutinin, it does not get rid of it, especially when cooked at lower temperatures, such as in a slow cooker. Phytohaemagglutinin can cross the gut barrier, increase intestinal permeability, and stimulate the immune system. In fact, phytohaemagglutinin is easily detected in the bloodstream after consumption. It has also been shown to cause extensive overgrowth of Escherichia coli, or E. coli, in the small intestine (some E. coli is normal, but it is a common culprit in small intestinal bacterial overgrowth). See also The Green Bean Controversy and Pea-Gate
Other agglutinins, like peanut agglutinin (PNA, or peanut lectin), also enter the bloodstream quickly after being consumed, implying significant effects on intestinal permeability. Agglutinins from nettle (typically consumed only cooked) increase intestinal permeability. Soybean agglutinin (soybean lectin) and concanavalin (an agglutinin in jack beans, which are often used in animal feeds) have been shown to increase epithelial permeability much as wheat germ agglutinin does. Concanavalin stimulates cytotoxic T cells and inflammatory cells from the innate immune system and is being investigated as a drug for chemotherapy (because chemotherapy drugs are more toxic to cancer cells than they are to normal cells, which seems to be the case with concanavalin). Soybean agglutinin is known to bind to and activate dendritic cells and stimulate proliferation of T cells. In fact, soybean agglutinin so efficiently targets and stimulates the immune system that it is being studied for use in vaccines to improve immunization.
These troublemakers are also found in vegetables from the nightshade family (see What Are Nightshades? and The WHYs behind the Autoimmune Protocol: Nightshades). The best understood nightshade lectin is tomato lectin, which is an agglutinin, more technically called lycopersicon esculentum agglutinin (LEA). (Try saying that five times fast!) Analogous to WGA, LEA binds to the surface of enterocytes and is known to cross the gut barrier. LEA has been investigated as an adjuvant (immune-stimulating chemical) for use in intranasal vaccines because it activates antibody production. An agglutinin from datura (a genus of plants belonging to the nightshade family, several of which are used as herbal medicines) also increases intestinal permeability.
Take-home message: even though wheat germ agglutinin has been most extensively studies, there’s plenty of evidence that other agglutinins, especially those from legumes and tomatoes, are equally as damaging.
Avoiding Agglutinins
Agglutinins cause a leaky gut and, through direct binding with immune cells, stimulate both the innate and adaptive immune systems. While some agglutinins (like phytohaemagglutinin) are largely deactivated by cooking, others (like wheat germ agglutinin) are not. Of course, when it comes to grains, pseudograins, legumes and nightshades, there’s other reasons to avoid them, discussed in the following posts:
- How Gluten (and other Prolamins) Damage the Gut
- Wheat and Innate Immunity
- Gluten-Free Diets Can Be Healthy for Kids
- The Link Between Gluten and Obesity
- Why Grains Are Bad: Part 1, Lectins and the Gut
- Why Grains Are Bad: Part 2, Omega 3 vs. 6 Fats
- Why Grains Are Bad: Part 3, Nutrient Density and Acidity
- How Do Grains, Legumes and Dairy Cause a Leaky Gut? Part 2: Saponins and Protease Inhibitors
- Are Pseudograins Pseudobad?
- The WHYs behind the Autoimmune Protocol: Nightshades
- 3 Myths About Legumes — Busted!
Trying to figure out how to avoid these foods? I have some handy dandy food lists in the following posts:
- How to Avoid Wheat and Gluten
- How to Avoid Soy
- How to Avoid Corn
- What Are Nightshades?
- Pseudograins: What Are They?
- Grains: What Are They?
- Legumes: What Are They?
- Paleo Diet Foods
Citations
Bardocz, S., et al., The effect of phytohaemagglutinin at different dietary concentrations on the growth, body composition and plasma insulin of the rat, Br J Nutr. 1996 Oct;76(4):613-26.
Dalla Pellegrina, C., et al., Plant lectins as carriers for oral drugs: is wheat germ agglutinin a suitable candidate? Toxicol. Appl. Pharmacol. 2005; 207, 170–178.
Dalla Pellegrina, C., et al., Studies on the joint cytotoxicity of wheat germ agglutinin and monensin, Toxicol. In Vitro. 2004; 18, 821–827.
Dowd, P.F., et al., Enhanced pest resistance of maize leaves expressing monocot crop plant-derived ribosome-inactivating protein and agglutinin, J Agric Food Chem. 2012 Oct 31;60(43):10768-75.
Greene, W.C., et al., Stimulation of immunoglobulin biosynthesis in human B cells by wheat germ agglutinin, J. Immunol. 1981; 127, 799–804
Greer, F. & Pusztai, A., Toxicity of kidney bean (Phaseolus vulgaris) in rats: changes in intestinal permeability, Digestion. 1985;32(1):42-6.
Gupta, R.K., et al., Soybean agglutinin coated PLA particles entrapping candidate vaccines induces enhanced primary and sustained secondary antibody response from single point immunization, Eur J Pharm Sci. 2012 Feb 14;45(3):282-95
Jordinson, M., et al., Systemic effect of peanut agglutinin following intravenous infusion into rats, Aliment Pharmacol Ther. 2000 Jun;14(6):835-40
Karlsson, A., Wheat germ agglutinin induces NADPH-Oxidase activity in human neutrophils by interaction with mobilizable receptors, Infect. Immun. 1999; 67,3461–3468
Kawakami, K., et al., Effect of wheat germ agglutinin on T lymphocyte activation, Microbiol. Immunol. 1988; 32, 413–422
Lochner, N., et al., Wheat germ agglutinin binds to the epidermal growth factor receptor of artificial Caco-2 membranes as detected by silver nanoparticle enhanced fluorescence, Pharm. Res. 2003; 20, 833–839
Ohba, H., et al., Cytoagglutination and cytotoxicity of wheat germ agglutinin isolectins against normal lymphocytes and cultured leukemia cell lines; relationship between structure and biological activity, Biochim. Biophys. Acta 2003; 1619, 144–150
Ohno, Y., et al., Effect of lectins on the transport of food ingredients in Caco-2 cell cultures, Biofactors. 2004;21(1-4):399-401
Pashkunova-Martic, I., et al., Lectin conjugates as biospecific contrast agents for MRI. Coupling of Lycopersicon esculentum agglutinin to linear water-soluble DTPA-loaded oligomers, Mol Imaging Biol. 2011 Jun;13(3):432-42
Pusztai, A., et al., Antinutritive effects of wheat-germ agglutinin and other N-acetylglucosamine-specific lectins, Br J Nutr. 1993 Jul;70(1):313-21
Reed, J.C., et al., Effect of wheat germ agglutinin on the interleukin pathway of human T lymphocyte activation, J. Immunol. 1985; 134, 314–323
Sjölander, A., et al., The effect of concanavalin A and wheat germ agglutinin on the ultrastructure and permeability of rat intestine. A possible model for an intestinal allergic reaction, Int Arch Allergy Appl Immunol. 1984;75(3):230-6.
Tang, S., et al., Effects of purified soybean agglutinin on growth and immune function in rats, Arch Anim Nutr. 2006 Oct;60(5):418-26
Watzl, B., et al., Dietary wheat germ agglutinin modulates ovalbumin-induced immune response in Brown Norway rats, Br. J. Nutr. 2001; 85, 483–490
Wirth, M., et al., Lectin-mediated drug delivery: discrimination between cytoadhesion and cytoinvasion and evidence for lysosomal accumulation of wheat germ agglutinin in the Caco-2 model, J. Drug Target. 2002; 10, 439–448


 TPV Podcast, Episode 313: The Bone Broth Show Part 2
TPV Podcast, Episode 313: The Bone Broth Show Part 2